

Research in Biology
Determining relative growth rates of bacterial isolates from marine biofilms
This work was conducted in the lab of Dr. Moisander, Department of Biology
By Andrea Pires

Andrea Pires at work in Dr. Moisander’s lab
Abstract
Biofilms of marine bacteria develop within hours of any surface submersed in seawater. Over the course of days and weeks these biofilms of bacteria mature to include protists, algae, cyanobacteria, and eventually unicellular and multicellular eukaryotes. Mature biofilms can interfere with marine operations such as boating and aquafarms, at which point they are referred to as biofouling. Antifouling methods have been developed to prevent biofouling, the most common of which involves coating the submersed surface with specialized toxic paint (de Carvalho, 2018). However, current antifouling methods are expensive and can be harmful to the environment. Potential targets for alternative antifouling methods could be found in the formation of biofilms at their early stages. There are many factors that are involved in early biofilm formation such as relative growth rates influencing bacterial fitness. This study focused on characterizing growth rates of 18 strains of marine bacteria recently isolated and identified from marine biofilms in Buzzards Bay. These cultures include bacteria from the Gammaproteobacterial genera Alteromonas, Pseudoalteromonas, Cobetia, Marinomonas, Salinimonas, Oceanobacter and Shewenella. The relative growth rates of the cultures were obtained through a series of 24-hour growth experiments. The estimated growth rates ranged from a minimum of 0.0847 h-1 to a maximum of 0.489 h-1. Significant differences in the growth rates among experiments were found for some strains. Significant differences were also found among the growth rates of strains within the genera Pseudoalteromonas, Salinimonas, and Shewenella but not among strains of Alteromonas sp.
Introduction
Biofilms are communities of adhered cells formed by bacteria and other microorganisms that cooperate to increase their chances of survival. In the marine environment, any submersed surface will rapidly develop a biofilm of bacteria that eventually matures to include other microorganisms such as protists, cyanobacteria and algae (de Carvalho, 2018). Advanced biofilms develop into biofouling, whose growth on submersed surfaces such as ship hulls and pipes can interfere with marine operations. For instance, the development of biofouling on ship hulls increases fuel consumption and fossil fuel emissions (Schultz et al., 2011). Various actions have been taken to eliminate biofouling, known as antifouling methods. The most common of these antifouling methods includes specialized toxic paint, which have their own negative impacts through toxicity to surrounding biota (de Carvalho, 2018). However, the costs and environmental impact associated with the development of biofouling and its prevention are substantial. Therefore, it is of interest to investigate the early stages of biofouling for purposes of exploring new potential targets for more affordable and environmentally sustainable antifouling methods.
In the initial stages of biofilm formation marine bacterial organisms colonize the submersed surface allowing for the adherence of successive colonizers such as protists and algae (de Carvalho, 2018). Targeting these initial bacterial colonizers could be an effective way to stop the development of biofouling at the source. However, more knowledge is needed about the growth dynamics of the initial biofouling colonizers. To begin, the relative growth
rates of 18 culture isolates of marine bacteria established and identified recently from marine biofilms in Buzzards Bay were investigated. All cultures used are common community members during early colonization of local marine biofilms (Naik et al., 2022). The methods and data from this experiment will help develop model systems for examining growth of initial biofouling colonizers and for designing further studies on impacts of antifouling methods on these bacteria.
Methods
Experimental Design
Reviving glycerol stocks
All culturing was conducted in a biosafety hood treated beforehand with ultraviolet light for at least 15 minutes. Sterile 5-mL tubes were used to revive the cultures from the glycerol stocks. The tubes were filled with 2.5-mL of marine broth. The glycerol stocks of the cultures were stored at -80ºC. The glycerol stocks of the cultures were kept on ice and using sterile, single-use inoculating loops, a small amount of the stock was obtained and transferred into its corresponding sterile 5-mL tube in the biosafety hood. The inoculated tubes were grown overnight (~24 hours) in a dark incubator at ~25-28ºC on a stirring plate.
Measuring Optical Densities (OD)
The next day, the optical densities of the overnight growth cultures were measured, and each strain was streaked on a Marine Agar plate (Fig. 1). 200 µL of each culture were added onto three wells of a non-treated polystyrene plate and read at 600 nm with a plate reader. These optical densities were then used to calculate the dilution factor for an OD of 0.05 and a total volume of 1000 µL. Using the subsequent dilution factors, a specific amount of each culture and Marine Broth were added to 2-mL sterile tubes. The optical densities of these dilutions were then measured again and recorded.
24-hour Growth Experiment
To prepare for the 24-hour growth experiment a plate lid was treated with 0.05% Triton X-100 in 20% ethanol to alleviate condensation forming in the lid throughout the 24-hour incubation. Each culture, adjusted to the approximate OD 0.05, further diluted with media to 1:100, was
aseptically pipetted to six replicate wells of a 96-well polystyrene plate. Each plate also included six wells with media only, serving as a negative control. The treated plate lid was then placed on top of the plate. The plate was then placed inside the microplate reader and grown at 25ºC. The plate reader program was adjusted to record optical density at 600 nm every 20 minutes for 24 hours. These readings were saved and used to estimate the growth rates of the marine bacterial cultures.
Estimating Growth Rates
The optical density readings recorded from the 24h growth experiments were then used to estimate the maximum growth rates of each strain and replicate (each well). The Easy Linear function within the R growthrates package was used to fit a linear model to the log- transformed data to find the maximum growth rate separately for each well, resulting in six separately calculated growth rates per strain per experiment (Petzoldt, 2020).
Results
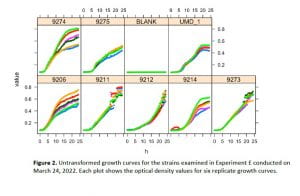
The 24h growth overnight experiment was performed for each of the bacterial cultures at least once and up to seven separate times. The untransformed growth curves obtained by the 24h experiments were plotted for each experiment using the R growthrates package (Fig. 2). The estimated maximum growth rates ranged from a minimum of 0.0847 h-1 to a maximum of 0.489 h-1 across all strains and replicates (Table 1).
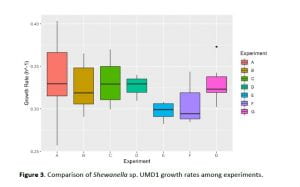
One of the aims of the experiments was to assess the methodological repeatability of the growth rate measurements across experiments. The variability in maximum growth rates of strains among experiments was analyzed using a series of one-way ANOVAs (Fig. 3). Growth rates of strains 9214 and 9273 were significantly different across experiments (one-way ANOVA, p < 0.05). Strains 9213, 9212, 9243 and 9275 were tested only once and the remaining strains showed no significant differences across experiments (one-way ANOVA, p>0.05). The growth rates of strains within the same genus were also compared using a series of one-way ANOVA tests. The Pseudoalteromonas genus included strains 9206, 9207 and 9274. There was a significant difference in the growth rates of strains 9206 and 9274 but other strains showed no differences. The Alteromonas genus included strains 9211, 9215, 9239, 9243, 9273, 9282, and 9283, and had no significant differences in their growth rates (p >0.05, Fig. 4). The Shewanella genus included strains 9242 and UMD1 with significantly different growth rates (p<0.05). The Salinimonas strains included strains 9275 and 9278 that also differed significantly in their growth rates.


Discussion
This study characterized growth rates of 18 recent marine biofilm bacterial isolates in the form of growth curves under standardized conditions. Calculated growth rates were used to examine any differences among the strains with respect to their growth patterns in monocultures. Variability across experiments is indicative of the reproducability and reliability of the growth rates obtained using this method. The data obtained from the growth experiments showed there was variability among experiments and strains within the same genus. However, only a few strains showed significant differences among experiments. For instance, strain 9213 showed significant differences in growth rates among experiments. This could have been influenced by the fact that it was later found to be a mixture of two different bacteria. Sources of variation within a single strain across experiments appear to have stemmed from periodically poor success in initial revival from glycerol or possibly experimental error.
Increasing the number of replicates per strain could help assess sources of variation among experiments.
There were significantly different growth rates among strains of the Pseudoalteromonas, Shewanella and Salinimonas genera but none were seen in the Alteromonas genus. These identified differences are likely due to slight functional differences among the strains but also could indicate more consistent differences among genera. Such differences may play a role in individual strain fitness in marine biofilms and will be of interest in future studies. Alteromonas spp. form a dominant component of early biofilms in local waters, while Shewanella spp. form a more subdominant group (Naik et al. 2022). A generally lower growth rate of Alteromonas was observed compared to Shewanella under the nutrient enriched conditions in these experiments and appears not to support the idea that representatives of Alteromonadaceae win during early biofilm colonization due to their superior growth rates.
Ultimately, these results will inform the growth curve methods and strains used in future experiments investigating biofilm formation along with competition and facilitation in co-cultures.
References
de Carvalho, C. C. C. R. (2018). Marine Biofilms: A Successful Microbial Strategy With Economic Implications. Frontiers in Marine Science, 5. https://www.frontiersin.org/article/10.3389/fmars.2018.00126
Naik, A., Smithers, M., & Moisander, P. H. (2022). Impacts of UV-C Irradiation on Marine Biofilm Community Succession. Applied and Environmental Microbiology, 88(4), e02298-21. https://doi.org/10.1128/aem.02298-21
Petzoldt, T. (2020). Estimation of Growth Rates with Package growthrates. https://tpetzoldt.github.io/growthrates/doc/Introduction.html
Schultz, M. P., Bendick, J. A., Holm, E. R., & Hertel, W. M. (2011). Economic impact of biofouling on a naval surface ship. Biofouling, 27(1), 87–98. https://doi.org/10.1080/08927014.2010.542809