Research in Physics
Characterizing the Friction Induced Triboelectric Effect of Polymers
By Viktoriya Balabanova

Portrait of Viktoriya at work in the lab
Introduction
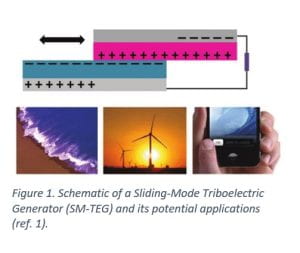
Triboelectric generators (TEGs) use the triboelectric effect between two layers of materials to convert mechanical movement (e.g. sliding) into electric power (Figure 1)1–4. These generators were developed in recent years and provide the most effective approach to harvest low-frequency mechanical energy for distributed energy applications. Up to now, almost all reported TEGs are designed to work in a charge-saturation state, meaning that more than enough force is used to guarantee maximum triboelectrification of materials in the device, which results in the waste of mechanical energy and low energy conversion efficiency.
Unlike previous studies, our research group hypothesizes that the TEGs will achieve maximum efficiency if they work under the minimum friction force that induces the saturation of triboelectric charges. This hypothesis cannot be tested until we know the friction force-triboelectric charge correlations in materials. This study aims to characterize such correlations in selected polymers. These polymers are the key materials that make the TEGs versatile, efficient, and cost-effective. The results of this project are significant because they will (1) provide preliminary data to test our significant hypothesis about TEGs, (2) help us gain new insights into previously unknown charge transfer mechanisms between two sliding surfaces.
Methods
We designed and built the setup shown in Figure 2, in order to measure the dynamic friction force and triboelectric charge induction simultaneously in a working sliding-mode triboelectric generator (SM-TEG). The whole setup consists of three major parts. Part 1 is a typical SM-TEG, consisted of two acrylic plates, on which adhesive spray is used to first attach stainless steel electrodes and then bind polymer sheets on top of them. Nylon film was used for the top plate (1×1 inch), and Polytetrafluoroethylene film (PTFE, 1×3 inch ) for the bottom. Part 2 of the setup consists of ATI-Nano17/IP68 six-axis force sensor firmly mounted and suspended from a frame. The top plate of the SM-TEG is attached to a specifically designed and 3-D printed holder, which is connected to the bottom of the force sensor. This second part of the setup remains fixed throughout the whole time data is being collected. In Part 3, a control mechanism is assembled for fine tuning of the normal force and the mechanical energy we put in the system. A very sensitive Newport M-270 Lab Jack is used to adjust the distance between the TEG’s plates, thus increasing or reducing it, allows us to control the normal force applied. For this part we hypothesize that the critical friction force that makes the system saturate can be very susceptible to changes, thus the precision adjustment of the normal force is required. On top of the lab jack a shaker plate PI C-891 is mounted and used to control the sliding mode of the TEG’s bottom plate, including amplitude, frequency and velocity. Another special holder for that plate was designed, 3-D printed and attached on top of the shaker. Lastly, the TEG’s electrodes are connected (open circuit) to a Keithley 6514 electrometer to read out the voltage/current induced by the triboelectric charges.

Results
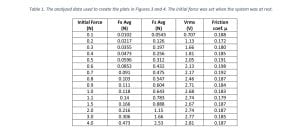
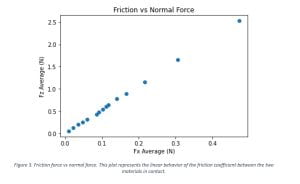
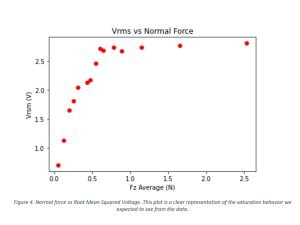
After the setup was built and calibrated, we initialized some preliminary tests to determine if the data from the force sensor and the electrometer was consistent with the behavior we expected to see. In this process we had to make micro adjustments of the setup, since we needed all parts of the TEG to be aligned and leveled. It was determined that we were ready to begin collecting data and we took several cycles of data sets for various initial forces. The obtained data was further analyzed and produced the results shown in Table 1, then it was plotted to create Figures 3 and 4.
Discussion
This summer research project aimed to characterize the correlation between friction force and triboelectric charge for selected polymers in pre-saturation states. The final results we obtained confirmed the behavior we expected to see. In Figure 3 the averages of the friction force and the applied normal force were analyzed and from the plot we can observe that the friction coefficient between the two polymers used in the TEG has a linear behavior. In Figure 4, the root-mean-squared voltage vs the normal force were examined. The charge was expected to increase with the applied force, until it reaches a saturation point under a critical force. This figure shows that after a certain threshold point, the increase of the normal force applied in the system does not increase the induced voltage. In other words, the TEG has reached a saturated state and even if we put more mechanical energy into it, it does not improve the overall performance of the TEG, resulting in low energy conversion efficiency.
This preliminary data is very important for us since it sets a clear direction for our future work. In the next year we are going to repeat the experiment and try to understand why this behavior happens, how the charge increases with force and how the critical force is affected by material properties. We are hoping to be able to create a model that will help improving the overall efficiency of TEGs, since they are a cost effective, easily manufactured, alternative source of green energy. I would like to thank Ross Jacques for his help on my project, and Dr. Caiwei Shen for his guidance and support, throughout the whole process, without whom this would not be possible!
References:
- Wang, Z. L. Triboelectric nanogenerators as new energy technology and self-powered sensors – Principles, problems and perspectives. Faraday Discuss. 176, 447–458 (2014). DOI:10.1039/C4FD00159A
- Xu, G., Li, X., Xia, X., Fu, J., Ding, W. & Zi, Y. On the force and energy conversion in triboelectric nanogenerators. Nano Energy 59, 154–161 (2019). DOI:10.1016/j.nanoen.2019.02.035
- Zhang, J., Darwish, N., Coote, M. L. & Ciampi, S. Static Electrification of Plastics under Friction: The Position of Engineering-Grade Polyethylene Terephthalate in the Triboelectric Series. Adv. Eng. Mater. 22, 1–5 (2020). DOI:10.1002/adem.201901201
- Rodrigues, C., Nunes, D., Clemente, D., Mathias, N., Correia, J. M., Rosa-Santos, P., Taveira-Pinto, F., Morais, T., Pereira, A. & Ventura, J. Emerging triboelectric nanogenerators for ocean wave energy harvesting: State of the art and future perspectives. Energy Environ. Sci. 13, 2657–2683 (2020). DOI:10.1039/d0ee01258k
- Zou, H., Guo, L., Xue, H., Zhang, Y., Shen, X., Liu, X., Wang, P., He, X., Dai, G., Jiang, P., Zheng, H., Zhang, B., Xu, C. & Wang, Z. L. Quantifying and understanding the triboelectric series of inorganic non-metallic materials. Nat. Commun. 11, 1–7 (2020). DOI:10.1038/s41467-020-15926-1



















 Portrait of Isabella Mancini, her colleagues, and mentor,
Portrait of Isabella Mancini, her colleagues, and mentor,